Among the various integumentary structures of vertebrates, feathers
are the most complex. Feathers are unique in their complex branching and
impressive variation in size, shape, color, and texture (Prum 1999,
Prum and Williamson 2001; Figure 1). Feathers were long considered the
defining anatomical feature of birds. However, many specimens of
non-avian dinosaurs have been discovered in China that show that
feathers are not restricted to birds (Figure 2). Specifically, most
non-avian coelurosaurian theropods appeared to have feathers, with
coelurosaurs being relatively small (2 – 3 m long), carnivorous
dinosaurs that occurred from the mid-Triassic through the early Jurassic
(230 – 200 million years ago).
Figure 1. The two basic feather types are pennaceous and plumulaceous
(or downy). Both types have a calamus. The pennaceous feather also has a
rachis (or shaft) from which branches the barbs. Branching off of the
barbs (upper right) are barbules. The hooklets of the barbules on the
distal side of barbs interlock with the barbules on the proximal side of
adjacent barbs. The ‘interlocked’ barbs on each side of the rachis form
the feather vanes. The plumulaceous feather has numerous
non-interlocked barbs extending from the calamus (From: Prum and Brush
2003).
In taxa more distantly related to birds, such as Sinosauropteryx
(Figure 3 below), multiple tufts projecting a few millimeters from the
skin have been discovered that resemble hypothesized early stages in
avian feather development. These filamentous ‘feathers’ (or
‘protofeathers’; there is some disagreement concerning whether or not
these integumentary structures were true feathers, e.g., Unwin 1998,
Lingham-Soliar et al. 2007) were about 20 (5-40) mm long and appear to
be rather homogenous over the body rather than originating in specific
tracts. To some investigators, the filaments appear to be like down
feathers and were probably used for insulation. They were hollow, and
appeared to have a short shaft with barbs, but no barbules. In 2009, a
fossil of another feathered dinosaur, Beipiaosaurus (a
coelurosaurian theropod), with even simpler feathers was reported (Xu et
al. 2009; Figures 4 and 5 below). These feathers consisted of single
broad (about 2 mm wide) filament, were 10 to 15 centimeters long, and
only present on the head, neck and tail. In taxa more closely related to
birds, such as the oviraptorid Caudipteryx and dromaeosaurid Sinornithosaurus,
elongate pinnate wing and tail feathers, structurally identical to the
feathers of present-day birds and comprised of a central rachis,
branching barbs, and barbules, have been found. In addition, fossils of a
Dromaeosaurid (Microraptor) have revealed asymmetrically veined pennaceous feathers on both the forelimbs and hindlimbs (Clarke and Middleton 2006).
Figure 3. Restoration of a Sinosauropteryx (Sinosauropteryx prima) with its body covered with feathers that were likely important for thermoregulation
(From: Chuong et al. 2001, and based on Chen et al. 1998).
Figure 4. The elongated, single filament feathers of Beipiaosaurus. The yellow arrows point to feathers on the head and neck (right) and tail (above)
(From: Xu et al. 2009).
Figure 5. Artist’s conception of Beipiaosaurus, a dinosaur with broad, single-filament feather (Image: Zhao Chuang and Xing Lida; source:
http://blogs.discovermagazine.com/80beats/2009/01/13/to-attract-mates-this-dino-may-have-shaken-a-tail-feather/).
Cladogram illustrating the relationship of birds with major groups of
non-avian coelurosaurian theropods. The numbers in circles at each
branching node indicate the first appearance of feathers and other key
morphological characters. 1, unbranched feathers; 2, uncinate processes on ribs; 3, true branched feathers; 4, retroverted pubis; 5, reversed hallux; 6, asymmetrical flight feathers;
7, pygostyle; 8, horny beak; 9, alula (bastard wing); 10, large, keeled
sternum. Taxa indicated with an asterisk are known to have possessed
either protofeathers or true feathers (From: Zhou et al. 2003).
Preserved evidence of archosaurian body covering
The earliest preserved scales,
filaments, or feathers are from the late Jurassic; the earliest crown
clade bird with feathers is from the Paleocene. Filamentous feather
precursors may have originated nearly 100 million years before the
origin of flight, but very few fossil deposits sample this period.
Sexual dimorphism in plumage and color patterning in Late Jurassic and
Early Cretaceous dinosaurs suggest that display functions played a key
role in the early evolution of pinnate feathers (Source: Clarke 2013).
Because birds evolved from reptiles and the integument of present-day
reptiles (and most extinct reptiles including most dinosaurs) is
characterized by scales, early hypotheses concerning the evolution of
feathers began with the assumption that feathers developed from scales,
with scales elongating, then growing fringed edges and, ultimately,
producing hooked and grooved barbules (Figure 6 below). The problem with
that scenario is that scales are basically flat folds of the integument
whereas feathers are tubular structures. A pennaceous feather becomes
‘flat’ only after emerging from a cylindrical sheath (Prum and Brush
2002). In addition, the type and distribution of protein (keratin) in
feathers and scales differ (Sawyer et al. 2000). The only feature shared
by feathers and scales is that they both begin development as a
morphologically distinct placode – an epidermal thickening above a
condensation, or congregation, of dermal cells (see Figure 8 below).
Feathers, then, are not derived from scales, but, rather, are
evolutionary novelties with numerous unique features, including the
feather follicle, tubular feather germ (an elevated area of epidermal
cells), and a complex branching structure (Prum and Brush 2002; Figure 7
below).
Figure 6. Hypothetical intermediate stage in the evolution of feathers
from scales, with ‘cracks’ separating sections of a large scales into
smaller, lateral plates, or protobarbs (From: Regal 1975).
Feathers are branched structures. The main branch of a typical
feather is the rachis, and barbs, consisting of a barb ramus and
projections called barbules, branch off the rachis (Figure above from
Prum and Brush 2003). Feathers grow from the base and the different
branches are generated by various mechanisms in the feather follicle.
Feather growth depends on nutrients provided via the follicular cavity
(dermal pulp), and the feather structure develops on the follicular (or
follicle) collar (inner epidermal layer; Figure 8 below). The production
of the complex branched structure involves the interaction of several
processes of cellular differentiation that occurs on the follicle collar
(Calcott 2009). Among present-day birds, variation in the shape and
structure of the rachis, barbs, and barbules generates a variety of
feather types, including flight (contour) feathers, semiplumes,
bristles, down feathers, filoplumes, and powder downs (Figure 7 below).
Figure 7. Various types of feathers of present-day birds, including
the contour feather (left) plus filoplumes, semiplumes, down feathers,
and bristles
(From: Lucas and Stettenheim 1972).
Figure 8. Schematic diagram of the development of a feather follicle.
(A) Development of the epidermal feather placode and the dermal
condensation. (B) Development of a feather papilla (or elongate feather
bud) via the proliferation of dermal cells. (C) Formation of the feather
follicle by the invagination of a cylinder of epidermal tissue around
the base of the feather papilla. (D) Cross-section of the feather
follicle as indicated by the dashed line in C. The follicle consists of a
series of tissue layers (from peripheral to central), including the
dermis of the follicle, the epidermis of the follicle (outer epidermal
layer), the follicle cavity or lumen (the space between epidermal
layers), the follicle (epidermal) collar (or inner epidermal layer), and
the dermal pulp (tissue at the center of the follicle). The
proliferation of feather keratinocytes and most of the growth of the
feather occurs in the follicle, or epidermal, collar (From: Prum 1999).
Pennaceous feather development
Downy feather development
Based on fossil evidence, we know that the first
non-avian theropods with simple, single-filament feathers lived about
190 million years ago, and that non-avian theropods with feathers having
a complex branching structure like those of present-day birds
(pennaceous feathers) existed about 135 million years ago. This fossil
evidence raises two important questions. First, if not derived from
scales, how did feathers evolve and, second, how did simple,
single-filament feathers evolve to become much more complex pennaceous
feathers? Of course, a related question is, given that non-avian
theropods did not fly, what function or functions did these feathers
serve?
Both fossil and developmental evidence suggests that feathers
evolved through a series of transitional stages, each the result of a
developmental evolutionary novelty or, in other words, a new mechanism
of growth (Prum 1999, Prum and Brush 2002, 2003). The first feathers,
like those of Beipiaosaurus , were unbranched, hollow
cylinders that developed from the tubular elongation (the feather germ)
of a placode (Figure 9 below). The advantage of a tubular feather germ
is that growth of a structure (in this case, a feather) can occur
without an increase in the size of the skin itself (in contrast to, for
example, scales; Prum 2005). An important step in the evolution of the
first feathers was a change in characteristics of the placode. Both
scales and feathers begin development from placodes, but feather
development, in contrast to scale development, requires generation of
suprabasal cell populations (dermal condensations) to form the follicle
(see Figure 8 above). The development of placodes where dermal
condensations occur, an evolutionary novelty, required changes in gene
expression and timing. However, such changes are known to be an
important mechanism in the origin of morphological innovations in many
other organisms (True and Carroll 2002, Prum 2005).
Figure 9. The first feathers were likely hollow cylinders (Stage I)
with undifferentiated collars that developed from an evolutionary novel
follicle collar
(From: Prum and Brush 2003).
Based on Prum’s (1999) model of feather evolution, the next step
after the origin of the feather follicle was the differentiation of the
follicle collar into barb ridges to generate barbs (Stage II; Figure 10
below). The resulting feather would consist of a tuft of barbs extending
from the calamus (Figure 10 below). Such a feather is hypothesized to
have evolved before the origin of the rachis (Stage IIIA) because the
rachis is initially formed by the fusion of barb ridges. In addition,
barbs are hypothesized to evolve before barbules because barbules
develop within layers of pre-existing barb ridges (Prum 1999). Feathers
comparable in structure to hypothesized Stage II feathers have been
reported from fossils of non-avian theropods, such as Sinornithosaurus mellenii (Figures 11 and 12 below; Xu et al. 2001, Norell and Xu 2005).
Figure 10. The next step in feather evolution (Stage II) involved the
differentiation of the follicle collar into barb ridges to generated
unbranched barbs (From: Prum and Brush 2003).
Figure 11. Filamentous integumental structure of Sinornithosaurus millenii
with compound structures composed of multiple filaments. These
structures exhibit two types of branching structure unique to avian
feathers: the filaments are joined in a basal tuft, and the filaments
are joined at their bases in series along a central filament. a, Arrows indicate the distal tips of some component filaments . b
, Illustrated reconstruction of the appendage showing the
positions of the observed filaments (lines) and the inferred outline of
the appendage (shading). Asterisk, the proximal end of the appendage.
The curved position of the appendage reveals its compound structure.
Each filament converges on the center of the appendage at its base.
Scale bar, 5 mm (From: Xu et al. 2001).
Figure 12. Sinornithosaurus millenii , based on this skeletal drawing by Marco Auditore and others
(Source: Wikipedia; http://en.wikipedia.org/wiki/File:Sinornithosaurus.jpg).
Line drawing of reconstructed skull of Sinornithosaurus. The skull is approximately 75 mm long.
vg, venom groove; mxf, maxillary fang; sff, subfenestral fossa; fc, fossa canal.
The birdlike raptor Sinornithosaurus was venomous -- Evidence suggests that some of the most avian dromaeosaurs, such as Sinornithosaurus,
were venomous. These raptors had unusual dentition and other cranial
features including grooved teeth, a possible pocket for venom glands,
and a groove leading from that pocket to the exposed bases of the teeth.
The anterior maxillary teeth are so long and fanglike that these
raptors appear to be saber-toothed. The long maxillary teeth do not
appear to have been deeply inserted
into the prey; the
grooved fang would probably have penetrated 4 to 6 mm into the skin.
This would be
sufficient to cut into the subdermal tissue and allow poison to enter
the bloodstream but would be too shallow to cause death or
immobilization through trauma alone. The poison of Sinornithosaurus
may have been similar in properties to rear-fanged snakes in that it
did not kill the envenomated animal quickly but rather placed it into a
rapid state of shock These features are all analogous to the venomous
morphology of lizards. Sinornithosaurus and related
dromaeosaurs probably fed on the abundant birds of the Jehol forests
during the early Cretaceous in northeastern China (Source: Gong et al. 2010).
The next step in feather evolution could have involved either the
development of a rachis via fusion of barbs or the development of
barbules that branched from the tufts of barbs. Perrichot et al. (2008)
discovered feathers from the Early Cretaceous (and preserved in amber;
Figure 15 below) that had shafts (rachis) consisting of incompletely
fused, still distinguishable, partially superimposed barbs. This
represents an intermediate stage between Prum’s (1999) stages II and
IIIa and suggests the possibility that rachis development may have
preceded barbule development (Figures 13 and 14 below).
Figure 13. Hypothesized stages I–III of feather evolution. Stage I of
this model assumes an unbranched, hollow filament, which developed from a
cylindrical invagination of the epidermis around a papilla. In stage
II, a tuft was formed by fusion of several filaments at their bases.
Stage III represents the formation of a central rachis and development
of serially fused barbs (III A) — to which, at a slightly later stage
(III B), secondary barbs (barbules) were added. The two other stages, IV
(bipinnate feathers with elaborate barbules and a closed vane) and V
(the asymmetrical flight feathers of modern flying birds), are not shown
(From: Sues 2001).
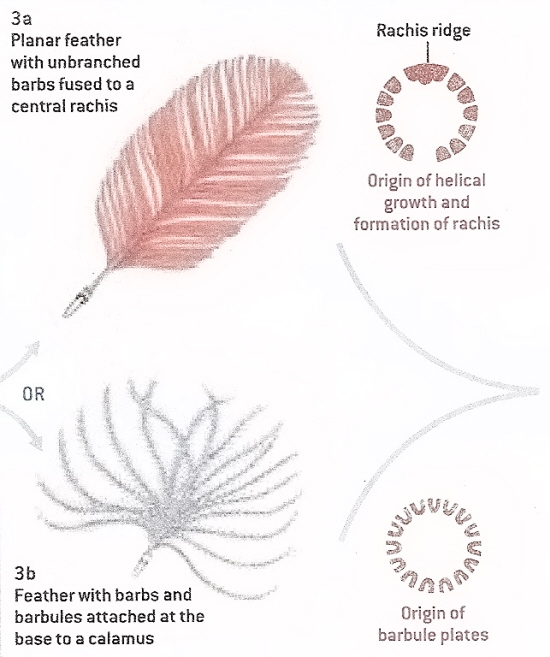
Figure 14. The next step in feather evolution (Stage
III) could have involved either the development of a rachis via fusion
of barbs (3a) or the development of barbules that branched from the
tufts of barbs (3b; From: Prum and Brush 2003). The discovery of
feathers from the Early Cretaceous that had shafts (rachis) consisting
of incompletely fused, still distinguishable, partially superimposed
barbs suggests that that rachis development (3a) may have preceded
barbule development (3b).
Figure 15. Three-dimensional virtual reconstruction of a fossil
feather from the Early Cretaceous (about 100 million years ago)
preserved in amber. This feather could be from either a bird or a
non-avian theropod. (a-c) long barbs form two vanes on each side of a relatively flattened shaft; (d)
the shaft is flattened and composed of incompletely fused bases of the
barbs, a stage in feather evolution that was hitherto unknown in fossil
records and corresponding to an intermediate stage between the very
distinct stages II and IIIa defined by Prum (1999). Scale bars, 100 µm
(From: Perrichot et al. 2008).
With the development of the rachis, the next stage in feather
evolution would likely have been the development of barbules (without
hooklets) to generate a bipinnate, open pennaceous structure (Stage 3a +
b; Figure 16 below). Subsequent evolution of differentiated proximal
and distal barbules would then generate the first closed, pennaceous
vane, with distal barbules growing hooklets to attach to the simpler,
grooved proximal barbules of the adjacent barb (Stage 4; Figure 16
below). Finally, lateral displacement of the new barb locus by
differential new barb ridge addition to each side of the follicle led to
the growth of a closed pennaceous feather with an asymmetrical vane
resembling modern remiges (Stage 5; Figure 16 below).
Figure 16. Hypothesized final stages in the evolution
of feathers like those of modern-day birds. T he development of
barbules (without hooklets) generated a bipinnate, open pennaceous
feather (Stage 3a + b), and evolution of differentiated proximal and
distal barbules led to the first closed, pennaceous vane, with some
barbules having hooklets to firmly attach to grooved barbules of the
adjacent barb (Stage 4). Differential new barb ridge addition to each
side of the follicle then led to the development of a closed pennaceous
feather with an asymmetrical vane (Stage 5) (From: Prum and Brush 2003).
Fossils reveal that the plumage sported by young Similicaudipteryx, especially on the forelimbs and tail,
was substantially different from that adorning older relatives (Credit: Xing Lida and Song Qijin).
Ontogenetic development of early feathers -- Recent
discoveries of feathered dinosaur specimens have greatly improved our
understanding of the origin and early evolution of feathers, but little
information is available on the ontogenetic development of early
feathers. Xu et al. (2010) described an early juvenile specimen and a
late-juvenile specimen, both referable to the oviraptorosaur Similicaudipteryx, recovered from the Lower Cretaceous (~125 million years ago) of western Liaoning, China.
The two specimens have strikingly different remiges and rectrices,
suggesting that a radical morphological change occurred during feather
development, as is the case for modern feathers. However,
both the remiges and the rectrices are proximally ribbon-like in the
younger specimen, but fully pennaceous in the older specimen, a pattern
not known in any modern bird. In combination with the wide
distribution of proximally ribbon-like pennaceous feathers and elongate
broad filamentous feathers among extinct theropods, this find suggests
that early feathers were developmentally more diverse than modern ones
and that some developmental features, and the resultant morphotypes,
have been lost in feather evolution.
Many dinosaurian groups, such as most ornithischians, the sauropodomorphs and the basal theropods,
are not included in this simplified dinosaurian cladogram. The
available specimens suggest that members of these groups had scaly skin,
but the possibility that they are partially covered by filamentous
integumentary structures cannot be completely excluded. Preservational
factors make it difficult to observe the detailed structure of the
filamentous feathers in available specimens of compsognathids, tyrannosauroids, and therizinosauroids,
so a ‘?’ is used to indicate uncertainty regarding the presence of
morphotypes 1, 3, 4 and 5 in these groups. On the basis of the
anatomical, ontogenetic, and phylogenetic distribution patterns of known
feather morphotypes among non-avian dinosaurs and early birds,
morphotypes 1, 2 and 7 are inferred to have been lost in feather
evolution, along with their associated developmental mechanisms.
Feather evolutionary-developmental model, terminology, and stage I and II specimens from Canadian amber. (A)
Current evolutionary-developmental model for feathers consists of a
stage I morphology characterized by a single filament: This unfurls into
a tuft of filaments (barbs) in stage II. In stage III, either some
tufted barbs coalesce to form a rachis (central shaft) (IIIa), or
barbules (segmented secondary branches) stem from the barbs (IIIb);
then, these features combine to produce tertiary branching (IIIa+b).
Barbules later differentiate along the length of each barb, producing
distal barbules with hooklets at each node to interlock adjacent barbs
and form a closed pennaceous (vaned) feather (stage IV). Stage V
encompasses a wide range of additional vane and subcomponent
specializations. Most modern birds possess stage IV or V feathers or
secondary reductions from these stages Green, calamus or equivalent;
blue, barbs; purple, rachis; red, barbule internodes; d.b., distal
barbules; r., ramus; p.b., proximal barbules. (B) Field of filaments cut obliquely (stage I). (C) Filament clusters variably oriented (stage II). (D)
Close-up of (C), showing filaments that comprise clusters. Pigmentation
coupled with comparatively thick outer walls produces darker color than
in isolated filaments. Scale bars, (B) and (C) 1 mm, and (D) 0.1 mm
(From: McKellar et al. 2011).
Late Cretaceous dinosaur and bird feathers in Canadian amber
-- The fossil record of early feathers has relied on carbonized
compressions that lack fine structural detail. Specimens in amber are
preserved in greater detail, but they are rare. Late Cretaceous
coal-rich strata from western Canada provide the richest and most
diverse Mesozoic feather assemblage yet reported from amber. The fossils
described by McKeller et al. (2011)
include primitive structures closely matching the protofeathers of
nonavian dinosaurs, offering new insights into their structure and
function. A Stage I feather (B in the figure above) contains a dense
forest of regularly spaced, flexible filaments; the closest
morphological match is the filamentous covering found of nonavian
dinosaurs such as
the compsognathid Sinosauropteryx prima. A Stage II morphotype
(C and D in the figure above) consists of tight clusters composed of
filaments; the most morphologically comparable
compression fossils are protofeathers associated with the dromaeosaurid Sinornithosaurus millenii.
Specialized barbules. (A) Coiled barbules surrounding thickened rachis (arrow), cut obliquely. (B) Close-up of coils in isolated barbule. (C)
Semi-flattened internodes and weakly expanded node of (A). Diffuse,
variable barbule pigmentation produces pale overall color. (D) Isolated barb with differentiated barbules and thickened ramus, in spider’s web. (E)
Barbules near distal tip of (D), with clearly defined distal and
proximal barbule series (left and right sides of ramus, respectively). (F)
Close-up of distal barbule in (E), showing nodal prongs and ventral
tooth on basal plate (arrow) adjacent to abrupt transition into
pennulum. Scale bars, (A) 0.4 mm; (B), (D), and (E) 0.2 mm; (C) and (F)
0.05 mm.
Other feathers have barbules specialized for discrete
functions. In A, B, and C above, a thickened rachis is surrounded by
numerous barbules with tightly coiled bases. The barbules undergo three
or more complete whorls and are composed of semi-flattened internodes
separated by weakly expanded node. Modern seedsnipes and sandgrouse
possess belly feathers with similar basal barbule coiling, allowing
water to be retained for transport to the nest for distribution to
nestlings or for cooling incubating eggs. Grebes also have coiled
barbules that absorb water into plumage, facilitating diving by
modifying buoyancy, reducing hydrodynamic turbulence, and improving
insulation. Barbules displaying all characteristics necessary for
forming vaned feathers are also present in Canadian amber (D, E, and F
above). These were probably borne by an animal capable of flight.
Barbules are widely spaced along a thick ramus (barb shaft) adapted for
rigidity and are strongly differentiated to interlock with adjacent
barbs to form a vane. On the basis of the presence of a rachis and
differentiated barbules, these feathers can be
assigned conservatively to stages IV and V and are attributed to Late
Cretaceous birds. These amber-preserved feathers demonstrate that
numerous evolutionary stages
were present in the Late Cretaceous, and that
plumage already served a range of functions in both dinosaurs and birds.
Early functional hypotheses for the origin of
feathers focused on their importance for flight (Steiner 1917, Heilmann
1926). However, the discovery of filamentous (and pennaceous) feathers
on flightless non-avian theropods provides clear evidence that feathers
evolved before the origin of flight and that the first feathers did not
serve an aerodynamic function. The earliest tuft-like feathers could
have served a variety of functions, including insulation, heat shielding
(Regal 1975), communication (Mayr 1960), crypsis (Prum 1999), water
repellency (Dyck 1985), and defense (Prum 1999).
The first cylindrical, filamentous feathers (Stage I) could have
provided insulation if they were sufficiently numerous. Feathers similar
in morphology to that predicted for Stage I feathers have been found on
fossils of Beipiaosaurus (a coelurosaurian theropod; Xu et
al. 2009). These primitive feathers consisted of single broad (about 2
mm wide) filaments, about 10 to 15 centimeters long, and were only
present on the head, neck and tail. Given their morphology and
distribution on the body, these feathers likely did not serve a
thermoregulatory function. Rather, their localized distribution and
morphology (relatively long and probably rather stiff) suggest that they
served as display structures (Xu et al. 2009). However, other types of
filamentous feathers in non-avian theropods more likely served a
thermoregulatory function (Norell and Xu 2005). For example, the
presence of dense filamentous feathers on Sinosauropteryx
suggests these theropods were endothermic, and that heat retention was
the primary function of the feathers (Chen et al. 1998; Figure 17
below).
Figure 17. a. Fossil of Sinosauropteryx prima. b
, Drawing of skeleton and feathers along the dorsal side and tail. Dark
pigmentation in the abdominal region might be some soft tissues of the
viscera (From: Chen et al. 1998).
The fossil of a pigeon-sized theropod, Epidexipteryx hui,
found in sediments from the Middle to Late Jurassic (152 - 168 million
years ago) of northern China revealed two pairs of elongate ribbon-like
tail feathers that probably served as ornaments (although they could
have also helped E. hui maintain balance when moving along tree
branches; Figure 18 below). These long feathers had a central shaft
(rachis) but, unlike the rectrices of present-day birds, the vanes were
not branched into individual filaments. Rather, they consisted of a
single ribbon-like sheet. Shorter feathers also covered the body and
could have served as insulation (Zhang et al. 2008). At present, Epidexipteryx is the oldest theropod known to have feathers that apparently served a display function.
Figure 18. Artists’ conception of Epidexipteryx hui showing
the paired ribbon-like tail feathers that likely served as ornaments and
played a role in intra- and intersexual interactions.
Illustration Credit: Zhao Chuang, Xing Lida/Nature.
Epidexipteryx hui
Pennaceous (or contour) feathers have been reported for a number of theropods, including the maniraptor Protarchaeopteryx (early Cretaceous; 120-122 million years ago), the oviraptorid Caudipteryx, and the dromaeosaurids Sinornithosaurus and Microraptor gui. Both Protarchaeopteryx and Caudipteryx had
pennaceous feathers (with barbules) on the forearms and tail (as well
as semiplumes and down-like feathers on the rest of the body). However,
the arms of these small theropods (about 0.4-0.7 kg) were relatively
short and all pennaceous feathers were symmetrical, indicating that
these dinosaurs could not fly or glide effectively. Some investigators
have suggested that these theropods, with relatively long legs and an
elevated hallux, were ground-dwelling runners (Qiang et al. 1998).
However, the forelimbs of Protarchaeopteryx and Caudipteryx,
although short relative to their hindlimbs, were longer than those of
other theropods and some investigators have argued that such elongation
(along with other characteristics, including recurved claws) suggests a
more (but not exclusive) arboreal lifestyle. For example, Chatterjee and
Templin (2004) argued that these theropods were largely arboreal and
that their small ‘protowings’ (in combination with the pennaceous
feathers on the tail) enhanced arboreal maneuvering and permitted
parachuting from branch to branch or from branch to ground (Figure 19
below). Feathers on the ‘protowings’ and tail would increase drag when
parachuting and, to some extent, slow the rate of descent, permitting a
safer landing. Another possible function of the forearm feathers is that
could have been used to increase hindlimb traction in the same manner
that some present-day birds, such as Chukar Partridges (Alectoris chukar),
flap their wings to improve hindlimb traction when they climb inclined
surfaces like the trunk of a tree (i.e., wing-assisted incline running;
Dial 2003, Clarke and Middleton 2006).
Figure 19. Although incapable of powered flight, the pennaceous feather of Protarchaeopteryx may have permitted parachuting, whereas those of Sinornithosaurus likely permitted gliding (From: Chatterjee and Templin 2004).
Other small theropods from the early Cretaceous (124-128 million years ago), including Sinornithosaurus and M. gui, had both plumulaceous and pennaceous feathers. Sinornithosaurus weighed about 1.5 kg, were likely arboreal, and, in contrast to Protarchaeopteryx and Caudipteryx,
their forelimbs were as long as their hindlimbs (Figure 12 above) . The
longer wings and greater wing surface area, in combination with
feathers on the tail (Figure 19 above), may have allowed Sinornithosaurus to glide between perches and from elevated perches to the ground (Chatterjee and Templin 2004).
M. gui was covered by plumulaceous
feathers about 25–30 mm long and feathers on the top of the head
were up to 40 mm long. Some feathers on the head were pennaceous
and probably served a display function. Large, asymmetric pennaceous
feathers were also attached to the distal tail, forelimb, and,
surprisingly, the hindlimb. Microraptor had both primary and
secondary flight feathers. This pattern was mirrored on the hind legs,
with flight feathers attached to the upper foot bones as well as the
upper and lower leg. When first described, Xu et al. (2003) proposed
that Microraptor was arboreal and glided from tree to tree with
four ‘wings’ – two forelimb wings and two hindlimb wings. However, Xu
et al. (2003) proposed that the legs extended out to the side (Figure 20
below) and other investigators pointed out that such a leg position was
unlikely because no known bird or theropod could extend their legs in
such a manner without dislocating the hip joint (Padian 2003).
Thereafter, Chatterjee and Templin (2007) proposed that the wings of Microraptor gui
would have been split-level (like a biplane) and not spread as
originally proposed, with the hindlimb flight feathers extending
horizontally and able to generate lift along with the forelimb wings
(Figure 21 below). Microraptor most likely employed a phugoid
(from the Greek, meaning take flight) style of gliding flight -
launching itself from a perch, swooping downward in a deep U-shaped
curve, and moving upward to land on a perch in another tree (Figure 21
below).
Figure 20. A reconstruction of Microraptor gui in gliding flight (Xu et al. 2003).
Gliding flight of the four-winged Microraptor gui -- Fossils of the remarkable dromaeosaurid Microraptor gui
and relatives clearly show well-developed flight feathers on the hind
limbs as well as the front limbs. No modern vertebrate has hind limbs
functioning as independent, fully developed wings; so, lacking a living
example, little agreement exists on the functional morphology or likely
flight configuration of the hindwing. Using a detailed reconstruction
based on the actual skeleton of one individual, cast in the round,
Alexander et al. (2010) developed light-weight, three-dimensional
physical models and performed glide tests with anatomically reasonable
hindwing configurations. Models were tested with hindwings abducted and
extended laterally, as well as with a previously described biplane
configuration. Although the hip joint requires the hindwing to have at
least 20° of negative dihedral (anhedral), all configurations were quite
stable gliders. Glide angles ranged from 3° to 21° with a mean
estimated equilibrium angle of 13.7°, giving a lift to drag ratio of
4.1:1 and a lift coefficient of 0.64. The abducted hindwing model’s
equilibrium glide speed corresponds to a glide speed in the living
animal of 10.6 m·s−1. Although the biplane model glided
almost as well as the other models, it was structurally deficient and
required an unlikely weight distribution (very heavy head) for stable
gliding. The model with laterally abducted hindwings represents a
biologically and aerodynamically reasonable configuration for this
four-winged gliding animal. M. gui’s feathered hindwings, although effective for gliding, would have seriously hampered terrestrial locomotion.
Microraptor preys on birds -- Preserved indicators of diet are extremely rare in the fossil record;
even more so is unequivocal direct evidence for predator–prey
relationships. O'Connor et al. (2011) report a unique specimen of the small
nonavian theropod Microraptor gui from the Early Cretaceous
Jehol biota, China, which has the remains of an adult enantiornithine
bird preserved in its abdomen, most likely not scavenged,
but captured and consumed by the dinosaur. This provides direct
evidence for the dietary preferences of Microraptor and a nonavian
dinosaur feeding on a bird. Further, because Jehol enantiornithines
were distinctly arboreal, in contrast to their cursorial ornithurine
counterparts, this fossil suggests that Microraptor may have hunted
in trees, supporting inferences that this taxon was also an
arborealist and providing further support for the arboreality of
basal dromaeosaurids.
Photograph and line drawing of a Microraptor gui fossil with remains of its avian prey shown in blue.
Figure 21. Top, teconstruction of M. gui in
dorsal view (left) showing the morphology and distribution of hindlimb
feathers and orientation of the hindlimb bones (above) during gliding.
Above right, cross-section showing relative position of forelimb and
hindlimb wings during gliding flight. Below right, a typical staggered
biplane (Stearman 75) for comparison with Microraptor; in biplane
aircraft of the 1920s, there was much additional drag generated by wires
and struts between the two wings, such drag-induced structures were
absent in Microraptor (Chatterjee and Templin 2007). Bottom,
hypothetical flight path showing a typical undulating phugoid path from
an initial take-off launch at a velocity of 3 meters per sec from a
perch 20-m high and landing safely at a speed of about 6.4 meters per
second (From: Chatterjee and Templin 2004).
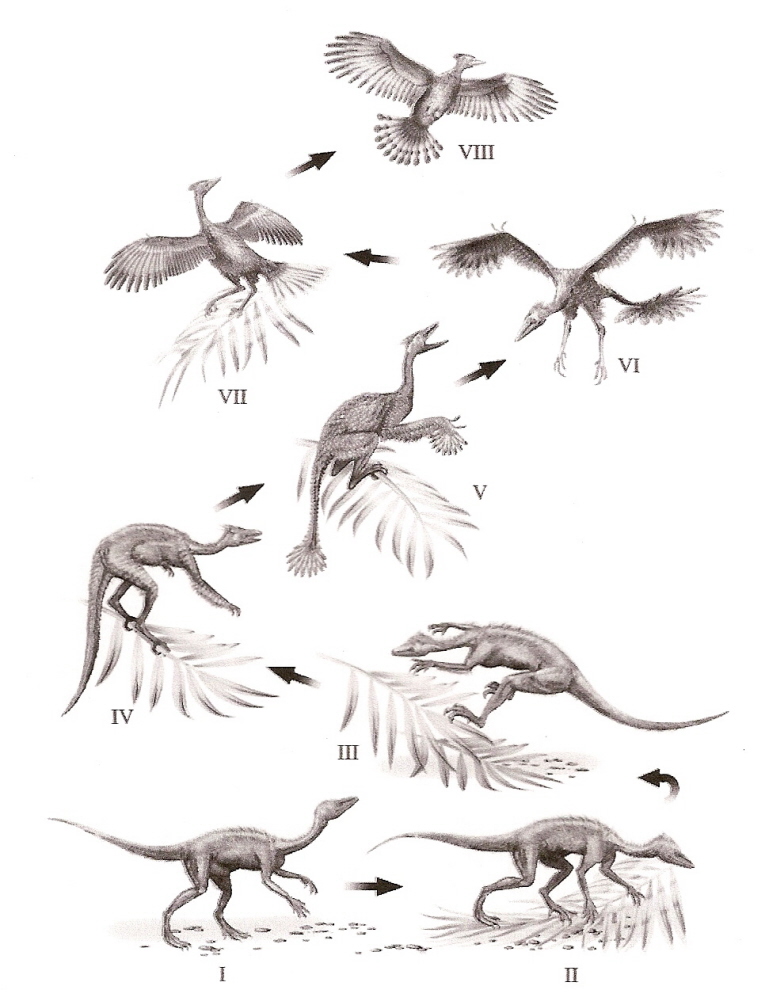
Possible scenario for the development of feathers leading to
the evolution of pennaceous feathers and flight. I and II, simple
feathers possibly important for thermoregulation or display; III, with
increasingly arboreal lifestyles, body feathers may have provided
ancestors of birds with a more aerodynamic shape useful for leaping
among branches; IV and V, simple pennaceous feather on the forelimbs may
have allowed parachuting; VI, larger pennaceous feathers with
symmetrical vanes may have permitted gliding; VII, asymmetrical feathers
may have contributed to more efficient gliding; VIII, powered flight
(From: Kurochkin and Bogdanovich 2008).
Current ideas about the evolution of feathers are based on dinosaurs (theropods) that actually lived well after Archaeopteryx.
Steps in the evolution of bird feathers were likely similar to those in
the evolution of feathers of non-avian theropods (Graphic source: PBS - NOVA).
Anchiornis huxleyi
A pre-Archaeopteryx troodontid theropod with feathers
-- The early evolution of the major groups of derived non-avialan
theropods is still not well understood, mainly because of their poor
fossil record in the Jurassic. A well-known result of this problem is
the ‘temporal paradox’ argument that is sometimes made against the
theropod hypothesis of avian origins (i.e., how could birds evolve from
theropods when the theropods seemingly most like those that gave rise to
birds actually existed after birds evolved). Hu et al. (2009) report on
an exceptionally well-preserved small theropod specimen collected from
the earliest Late Jurassic Tiaojishan Formation (151 - 161 million years
old) of western Liaoning, China. The specimen is referable to the
Troodontidae, which are among the theropods most closely related to
birds. This new find refutes the ‘temporal paradox’ because Anchiornis
lived several million years before Archaeopteryx and provides
significant information on the temporal framework of theropod
divergence. Furthermore, the extensive feathering of this specimen,
particularly the attachment of long pennaceous feathers to the pes
(legs), sheds new light on the early evolution of feathers and
demonstrates the complex distribution of skeletal and integumentary
features close to the dinosaur–bird transition.
The long pennaceous leg feathers of Anchiornis, like those previously
described for Microraptor and Pedopenna and Anchiornis, indicate that
they served an aerodynamic function. This would imply that a four-winged
condition played a role in the origin of avian flight and, because long
feathers on the legs would likely impede terrestrial locomotion, also
lends support to the arboreal hypothesis for the development of flight.
Calcott, B. 2009. Lineage explanations: explaining how biological
mechanisms change. British Journal for the Philosophy of Science, online
early.
Chatterjee, S., and R. J. Templin. 2004. Feathered coelurosaurs from
China: new light on the arboreal origin of avian flight. In: Feathered
dragons (P. J. Currie, E. B. Koppelhus, M. A. Shugar, and J. L. Wright,
eds.), pp. 251-281. Indiana University Press, Bloomington, IN.
Chatterjee, S., and R. J. Templin. 2007. Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. Proceedings of the National Academy of Science 104: 1576-1580.
Chen, P., Z. Dong, and S. Zhen. 1998. An exceptionally well-preserved
theropod dinosaur from the Yixian Formation of China. Nature 391:
147-152.
Chuong, C. M., L. Hou, P. J. Chen, P. Wu, N. Patel, and Y. Chen.
2001. Dinosaur’s feather and chicken’s tooth? Tissue engineering of the
integument. European Journal of Dermatology 11: 286-292.
Kurochkin, E. N., and I. A. Bogdanovich. 2008. On the origin of avian
flight: compromise and system approaches. Biology Bulletin 35: 1-11.
Lingham-Soliar, T., A. Feduccia, and X. Wang. 2007. A new Chinese
specimen indicates that 'protofeathers' in the Early Cretaceous theropod
dinosaur Sinosauropteryx are degraded collagen fibres. Proceedings of the Royal Society B 274: 1823-1829.
Lucas, A. M., and P. R. Stettenheim. 1972. Avian anatomy. Integument.
Agriculture Handbook 362, U.S. Department of Agriculture, Washington,
D.C.
Mayr, E. 1960. The emergence of evolutionary novelties. In: The
evolution of life (S. Tax, ed.), pp. 349-380. University of Chicago
Press, Chicago, IL.
Padian, K. 2003. Four-winged dinosaurs, bird precursors, or neither? BioScience 53: 451-453.
Perrichot, V., L. Marion, D. Néraudeau, R. Vullo, and P. Tafforeau.
2008. The early evolution of feathers: fossil evidence from Cretaceous
amber of France. Proceedings of the Royal Society B 275: 1197-1202.
Prum, R. O. 1999. Development and evolutionary origin of feathers. Journal of Experimental Zoology 285: 291-306.
Prum, R. O. 2005. Evolution of the morphological innovations of feathers. Journal of Experimental Zoology 304B: 570-579.
Prum, R. O., and A. H. Brush. 2002. The evolutionary origin and
diversification of feathers. Quarterly Review of Biology 77: 261-295.
Prum, R. O., and A. H. Brush. 2003. Which came first, the feather or the bird? Scientific American 286: 84-93.
Prum, R. O., and S. Williamson. 2001. Theory of the growth and
evolution of feather shape. Journal of Experimental Zoology 291: 30-57.
Qiang, J., P. J. Currie, M. A. Norell, and J. Shu-An. 1998. Two
feathered dinosaurs from northeastern China. Nature 393: 753-761.
Regal, P. J. 1975. The evolutionary origin of feathers. Quarterly Review of Biology 50: 35-66.
Sawyer, R. H., T. Glenn, J. O. French, B. Mays, R. B. Shames, G. L.
Barnes, Jr., W. Rhodes, and Y. Ishikawa. 2000. The expression of
beta-keratins in the epidermal appendages of reptiles and birds.
American Zoologist 40: 530-539.
Steiner, H. 1917. Das problem der diastataxie des vogelflügels. Jenaische Zeitschrift für Naturwissenschaft 55: 222-496.
True, J. R., and S. B. Carroll. 2002. Gene co-option in physiological
and morphological evolution. Annual Review of Cell and Developmental
Biology 18: 53-80.
Unwin, D. M. 1998. Feathers, filaments and theropod dinosaurs. Nature 391: 119-120.
Xu, X., X. Zheng, and H. You. 2009. A new feather type in a nonavian
theropod and the early evolution of feathers. Proceedings of the
National Academy of Science USA 106: 832-834.
Xu, X., Z. Zhou, and R. O. Prum. 2001. Branched integumentary structures in Sinornithosaurus and the origin of feathers. Nature 410: 200-204.
Xu, X., Z. Zhou, X. Wang, X. Kuang, F. Zhang, and X. Du. 2003. Four-winged dinosaurs from China. Nature 421: 335-340.
Zhang, F., Z. Zhou, X. Xu, X. Wang, and C. Sullivan. 2008. A bizarre
Jurassic maniraptoran from China with elongate ribbon-like feathers.
Nature 455: 1105-1108.
Zhou, Z., P. M. Barrett, and J. Hilton. 2003. An exceptionally preserved Lower Cretaceous ecosystem. Nature 421: 807-814.